2.4 Animal eyes⧉
Human vision is one solution among many, and a parochial one. Eyes have evolved independently dozens of times, and surveying the alternatives is the best cure for treating our own visual system as the definition of "seeing." Two questions organize the tour, the same two that organize the whole book's treatment of a camera: how is the image formed (optics), and how is color sensed (the photoreceptors and their wiring). The optical side connects back to Image formation; the color side extends the opsin story of Perceptual color and trichromatic vision.
2.4.1 The optics: many ways to form an image⧉
Forming an image means getting light from each direction in the scene onto a different receptor — the same plenoptic-sampling problem a camera solves with a lens. Nature has found several radically different answers (Figure 2.4.2).
Before the variety, the sequence. A full eye is not an all-or-nothing leap but a ladder of small, individually useful improvements (Land & Nilsson Land & Nilsson, Animal Eyes). Start with a flat patch of photoreceptors that registers only light versus dark; fold it into a cup, and the rim shades part of the sheet, giving a crude sense of direction; close the cup to a pinhole, and a real (if dim) inverted image forms with no lens at all; finally fill the aperture with a lens for a bright, focused image. That progression — from no image to a focused one — recapitulates the optics of this book in miniature (Figure 2.4.1).

- Single-lens ("camera") eyes, like ours and the octopus's, use one refracting element to cast a single inverted image on a sheet of receptors — the design we have been modeling all chapter. The vertebrate and cephalopod versions evolved separately and even wire their retinas back-to-front relative to each other, a textbook case of convergent evolution onto the same optics.
- Compound eyes (insects, crustaceans) tile the visual field with hundreds or thousands of ommatidia, each a tiny lens feeding one or a few receptors and pointing in a slightly different direction. Resolution is set by how finely the facets are packed, not by a single aperture — so acuity is modest, but the field of view is enormous and the temporal response very fast (one reason flies are so hard to swat). It is, in effect, a fixed array of one-pixel cameras.
- Lens-less designs solve image formation without refraction at all: the pinhole eye of the nautilus (an aperture with no lens — sharp but dim, exactly the pinhole-camera tradeoff from Image formation), and the mirror ("reflecting") eyes of scallops and some deep-sea crustaceans, which focus by a curved multilayer mirror behind the retina rather than a lens — the biological analogue of a catadioptric camera.
- Eyes built for a niche. Many designs are tuned to a task more than to general acuity: the tapetum lucidum, a reflective layer behind the retina in cats, dogs, and deer, bounces unabsorbed photons back through the receptors for a second chance — boosting night sensitivity at the cost of a little blur (and causing eyeshine in a flash photo). Deep-sea fish stack receptors or grow tubular eyes to wring signal from a few photons; the four-eyed fish Anableps splits its cornea to focus above and below the waterline at once.


2.4.2 The color vision: opsins remixed⧉
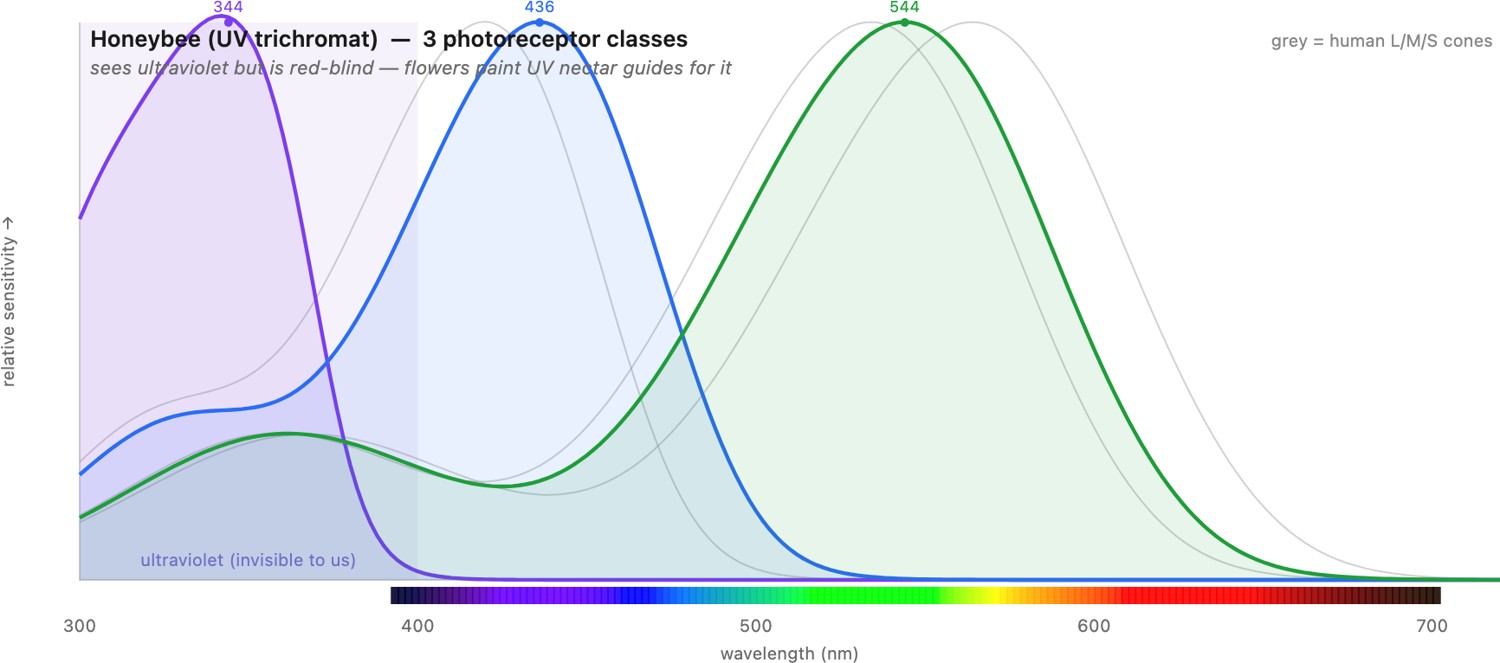
If the optics vary, color vision varies even more, because "color" is nothing but whichever set of opsins (the light-sensitive photopigment proteins) an animal happens to carry — and, crucially, how it compares their outputs. Laid side by side, the photoreceptor spectral sensitivities of different animals make the diversity concrete (Figure 2.4.4): where we have three heavily-overlapping cones, a bee has three shifted into the ultraviolet, a bird or a fish has four, a dog has two, and a mantis shrimp has a dozen narrow ones spanning deep UV to red. Opsins are not the whole story: many birds, reptiles, and fish place colored oil droplets in front of their photoreceptors, tiny filters that absorb part of the incoming spectrum before it reaches the pigment. This lets them sharpen and shift their effective spectral sensitivities away from the broad opsin curves, building narrower, better-separated color channels than the pigments alone would give. Three lessons stand out, all of them sharpening points made in Perceptual color and trichromatic vision.

- More dimensions than three. Our trichromacy is not a ceiling. Many birds, reptiles, and fish are tetrachromats, with a fourth cone reaching into the ultraviolet (UV) — they see patterns on flowers and plumage that are simply invisible to us. Most mammals, by contrast, are dichromats (red–green "color-blind" by our standard); primate trichromacy is re-evolved, the product of a recent gene duplication that split one pigment into our overlapping L and M cones — which is exactly why L and M overlap so heavily (see the opsin gene tree below). The lesson from the metamerism section of Perceptual color and trichromatic vision returns with teeth: every species is "color-blind" relative to some other, ourselves included, because every visual system projects an infinite spectrum onto a handful of numbers.

- More receptors do not mean more discrimination. The mantis shrimp carries roughly twelve to sixteen photoreceptor classes — and yet discriminates colors rather poorly in behavioural tests Thoen et al. 2014. The leading interpretation is that it does not compare channels the way a trichromat does (the comparison that the univariance discussion in Perceptual color and trichromatic vision showed is essential); instead it appears to recognize colors almost directly, reading off which receptor fires hardest. It is a working counterexample to "more channels = better color," and a reminder that color lives in the wiring, not just the pigments.
- Sampling axes we lack entirely. Some animals sense components of the plenoptic function we cannot. Many insects and cephalopods are sensitive to the polarization of light, using it to navigate by the sky's polarization pattern or to cut glare off water (recall polarization from Light and physics) — the mantis shrimp even senses circular polarization. Others extend their spectral window well outside our 400–700 nm band: pit vipers image thermal infrared with a separate pit organ, and UV-sensitive species push the short-wavelength end. Each species samples a different slice of the same physical world.
All of this divergence is written in the opsin gene tree. From a single ancestral opsin, gene duplication and loss produced the pigment classes that different animals draw on, and our own re-evolved L/M split is one recent twig on it — which is why those two cones overlap so heavily (Figure 2.4.6). An interactive explorer lets you dial up any species' photoreceptor curves against our own and watch its world of color reach past ours (Figure 2.4.7). The phylogeny is developed in the "Evolution of Eyes" chapter of Vision (Cambridge) Vision, "Evolution of Eyes".

The throughline of the whole chapter — now stated across species: color and vision are not read off the world, they are computed from it — projected through whatever opsins an eye carries, re-coded by whatever wiring sits behind them, adapted, and inferred. Human vision is one particular such computation; every later chapter on color, encoding, and image quality is an attempt to live within, or exploit, the constraints our version of that biological processor imposes.