2.2 Perceptual color and trichromatic vision⧉
Color has several meanings from physical to different layers of perception and all these stages matter to photography and can be the cause of much confusion. This chapter follows that whole pipeline in order: from physical color (a spectrum), to three cone responses, to the opponent recoding, up to the high-level assessments where illuminant discounting, color constancy, color names, and focal prototypes live. Each stage is a different, equally valid answer to "what color is this." The rest of human vision — light adaptation, lightness and color constancy, contrast, spatial and temporal sensitivity, and attention — is taken up in the next chapter, Human Vision.
One point is worth flagging: visual perception is measurable. Psychophysics, the experimental study of the map from stimulus to sensation, is quantitative and reproducible. Color-matching, hue-cancellation, and contrast-sensitivity experiments return tight numbers that repeat across observers and across laboratories, solid enough to be frozen into standards such as the CIE standard observer and the perceptual models inside every camera and codec.
2.2.1 Visual system anatomy:⧉
Light entering the eye passes through the cornea and the lens, the eye's two refracting elements, the same Snell's-law optics as a camera lens (the cornea does most of the bending; the lens fine-tunes focus by changing shape, accommodation). The pupil, the iris's adjustable opening, is the eye's aperture. The image lands on the retina, a thin sheet of neural tissue lining the back of the eyeball (Figure 2.2.1).
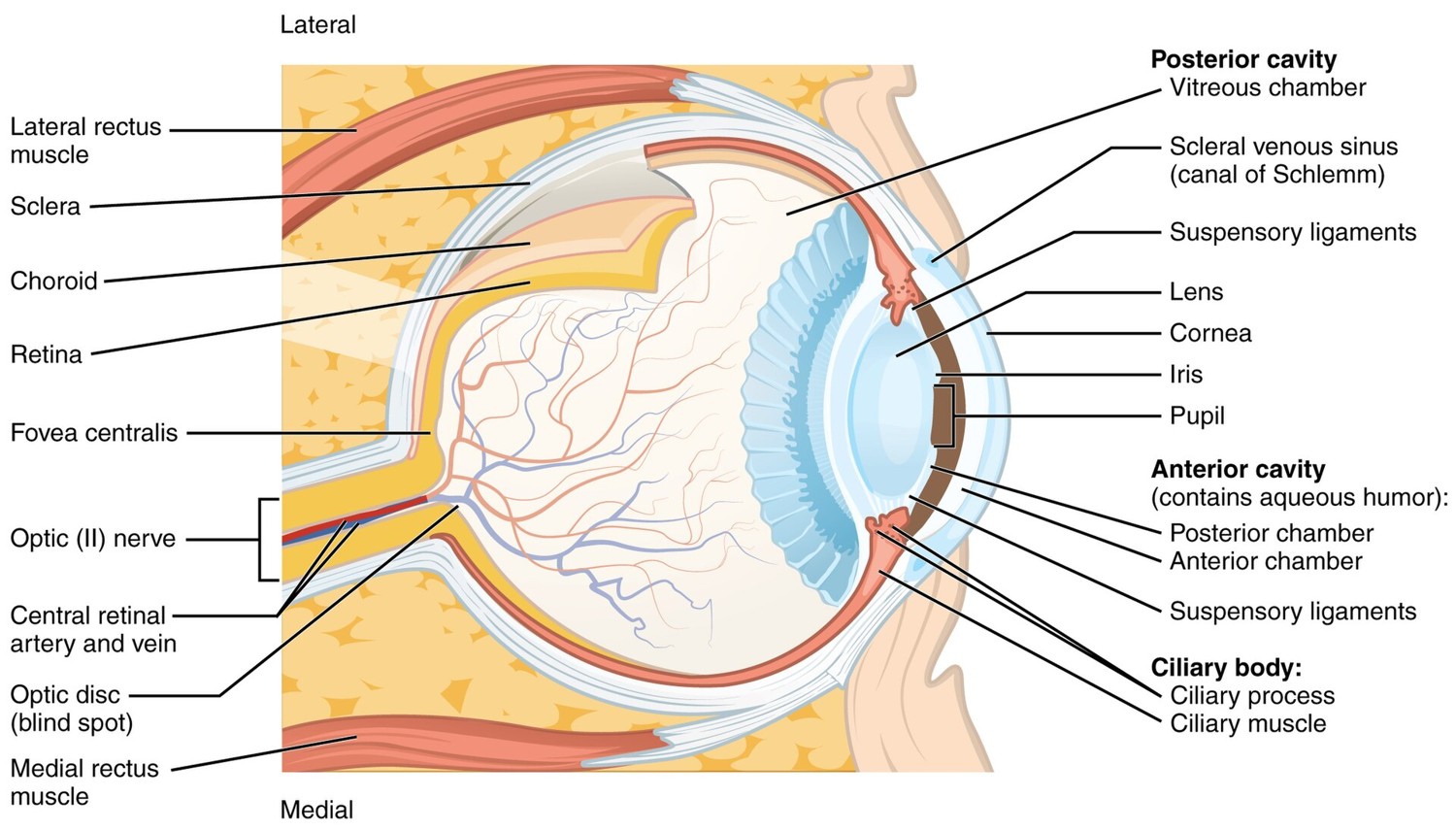
The retina's light-sensitive cells come in two families. Rods are exquisitely sensitive and handle dim, night (scotopic) vision, but there is only one kind, so rod vision is colorless. Cones are less sensitive, work in daylight (photopic vision), and come in three types — the basis of color. Their distribution across the retina is wildly non-uniform (Figure 2.2.2). Cones are packed densely in the fovea, the tiny central pit we point at whatever we are scrutinizing; rods dominate the periphery. This is why you cannot read text out of the corner of your eye, and why faint stars are easier to see slightly off-center.
Physically these are minute cells. In the fovea the cones are the finest of all, only about $2\,\mu\text{m}$ across, packed at a center-to-center spacing near $2.5\,\mu\text{m}$ that sets the eye's acuity limit; toward the periphery cones fatten to $5$–$8\,\mu\text{m}$. Rods are slimmer still, roughly $1$–$2\,\mu\text{m}$ in diameter. A few micrometers is therefore the biological yardstick against which a camera's photosite is measured, and modern phone pixels have shrunk to below the size of a foveal cone (→ photosite sizes).


Two quirks of this layout matter. First, there are essentially no short-wavelength (S, "blue") cones in the very center of the fovea, and far fewer S cones than long- and medium-wavelength (L and M) cones overall. As a result blue is sensed at low resolution and is even slightly out of focus — the eye's chromatic aberration puts blue at a different focal plane — yet we perceive a crisp blue everywhere, because the brain fills it in. Second, luminance (our sense of brightness) is roughly the sum of the L and M cone responses, so green carries most of our spatial acuity. This single fact echoes through the whole book: it is why camera color-filter mosaics (the Bayer pattern) use twice as many green cells as red or blue, and why image compression can throw away color detail but not brightness detail.
The retina is not a passive film: it does substantial processing before anything leaves the eye. The photoreceptors do not report straight to the brain; they feed layers of intermediate cells that in turn drive the retina's output neurons, the ganglion cells, whose long fibers bundle together into the optic nerve, the cable to the brain. Already here the signal is reorganized: each output cell no longer reports raw brightness but compares a small spot to the ring of retina around it, a center-surround response that reacts to local contrast and edges rather than absolute level (the seed of the edge enhancement we return to under Spatial vision). From the eye the pathway passes through a relay station deep in the brain (the lateral geniculate nucleus) and on to the primary visual cortex at the back of the head, then to higher areas that handle motion, form, and recognition. The lesson for us: much of what we naïvely attribute to "the image" is really a construction, assembled in stages from a heavily pre-processed retinal signal.
2.2.2 Color⧉
Even though cones are known to be the color vision receptors, a single cone does not measure wavelength — it produces one number, and stimuli of different wavelengths can yield the same response.
A given cone type $k$ has a spectral sensitivity $c_k(\lambda)$: a curve saying how strongly it responds to light of each wavelength (Figure 2.2.4).

When light with spectrum $E(\lambda)$ arrives, the cone reports a single response, the spectrum weighted by its sensitivity and summed over all wavelengths (Figure 2.2.5):
Mathematically this is a dot product. In the case of monochromatic light (a single wavelength $\lambda$) it reduces to the scalar product of the cone's sensitivity at $\lambda$ and the light's intensity for $\lambda$.
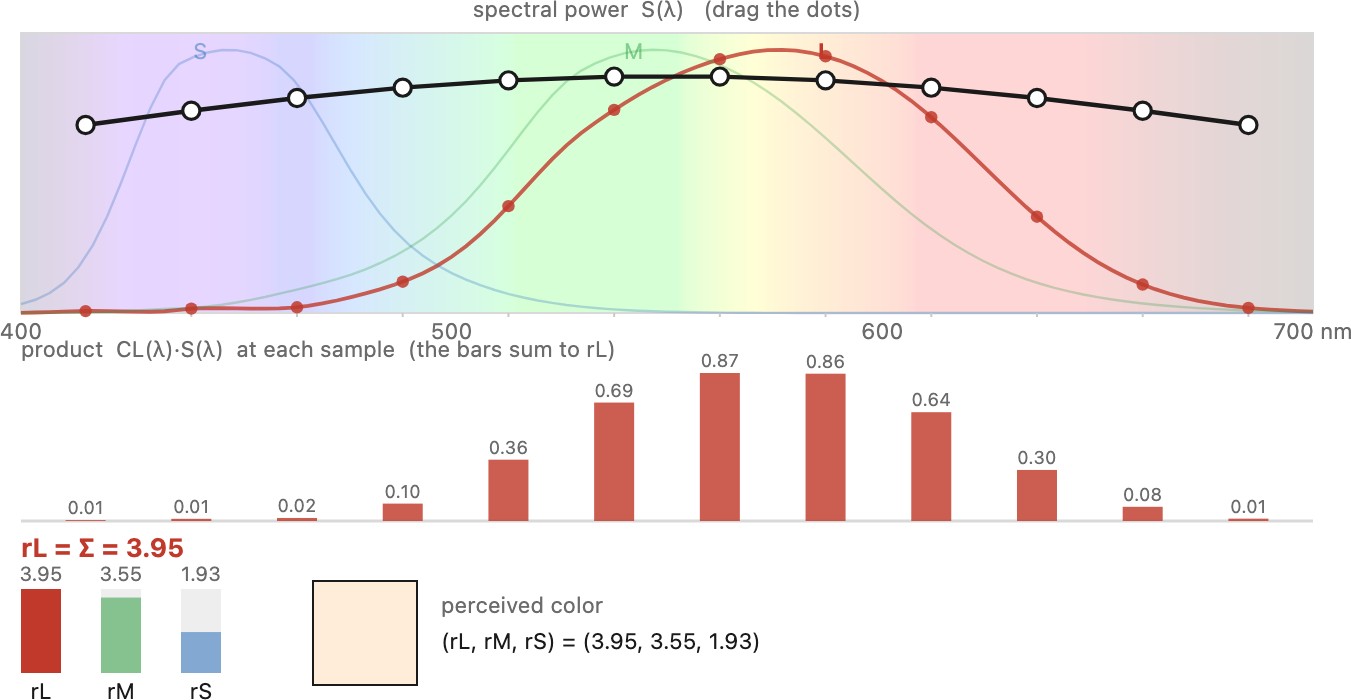
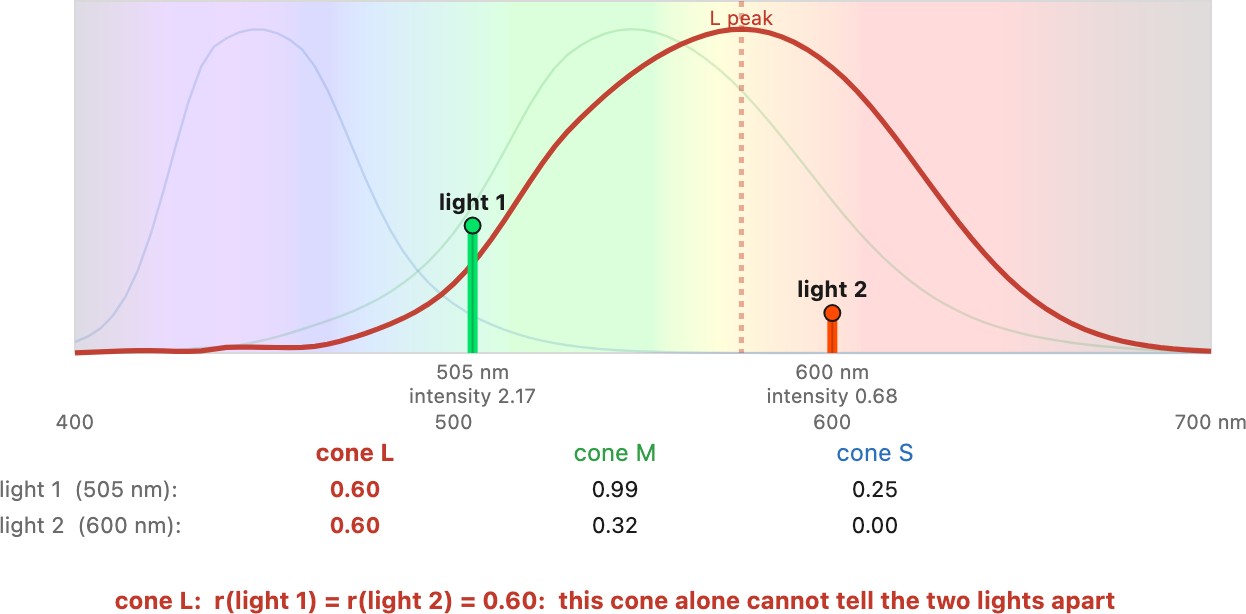
As Figure 2.2.6 shows, lights at different wavelengths can yield the same response if their intensity ratio is the inverse of the cone's spectral response ratio for those two wavelengths. A cone conflates color with intensity.
While a single cone cannot distinguish wavelength, the combination of three cone responses provides much more information (click on "show other cones" in Figure 2.2.6 ). This is the trichromatic theory of color vision, pioneered by Thomas Young and put on a quantitative footing by Hermann von Helmholtz. Three cone types give three numbers $(r_L, r_M, r_S)$, and that triple is all the information the eye keeps about the spectrum. An infinity of wavelengths collapses to three.
Hermann von Helmholtz (1821–1894) was the physicist and physiologist who turned Thomas Young's guess into a working theory. Young had argued decades earlier that the eye cannot possibly carry a separate detector for every wavelength and must instead make do with a few receptor types; Helmholtz developed this quantitatively, and the three-receptor account of color vision is still called the Young–Helmholtz theory. His Handbook of Physiological Optics founded the field, and he invented the ophthalmoscope that let physicians look into the living eye. Two of his ideas run straight through this chapter. The first is trichromacy itself, the L/M/S projection that everything here is built on. The second is unconscious inference, his claim that perception is not a direct readout of the retina but an inference the brain makes about the most likely cause of its input. That is exactly the stance we take later on color constancy, white balance, and "the dress," where the visual system guesses the illuminant and quietly discounts it. Beyond vision he is everywhere in physics as well, from the conservation of energy to the Helmholtz free energy and the acoustic resonators that carry his name.
It helps to make the projection concrete as linear algebra. Discretize the spectrum into $N$ wavelength bins and stack the samples into a vector $\mathbf{E} \in \mathbb{R}^N$. Stack the three sensitivity curves as the rows of a $3 \times N$ matrix $\mathbf{C}$, one row per cone type. Then the three cone responses are just a matrix–vector product,
(Figure 2.2.7). In reality wavelength is continuous, so $N \to \infty$ and the sum becomes the integral above — color sensing is genuinely an infinite-dimensional input squeezed through a three-dimensional bottleneck.

Each of the three cone types reports one number: the integral over wavelength of the incoming spectrum times that cone's spectral sensitivity — a projection (dot product) of the spectrum onto the cone curve, $r_k=\sum_i c_k(\lambda_i)\,E(\lambda_i)=\mathbf c_k\!\cdot\mathbf E$. Three cones → three dot products → the entire input to human color. Everything else about color — metamerism, "primary" colors, the color-matching matrices of the next chapter — follows from this one fact: color is three dot products with three fixed curves.
We established above that a single cone cannot distinguish wavelengths. Only by comparing the three cones' outputs can the brain begin to separate the two. However, this ability is limited. This is unsurprising because trichromatic vision summarizes a while spectrum with only three numbers.
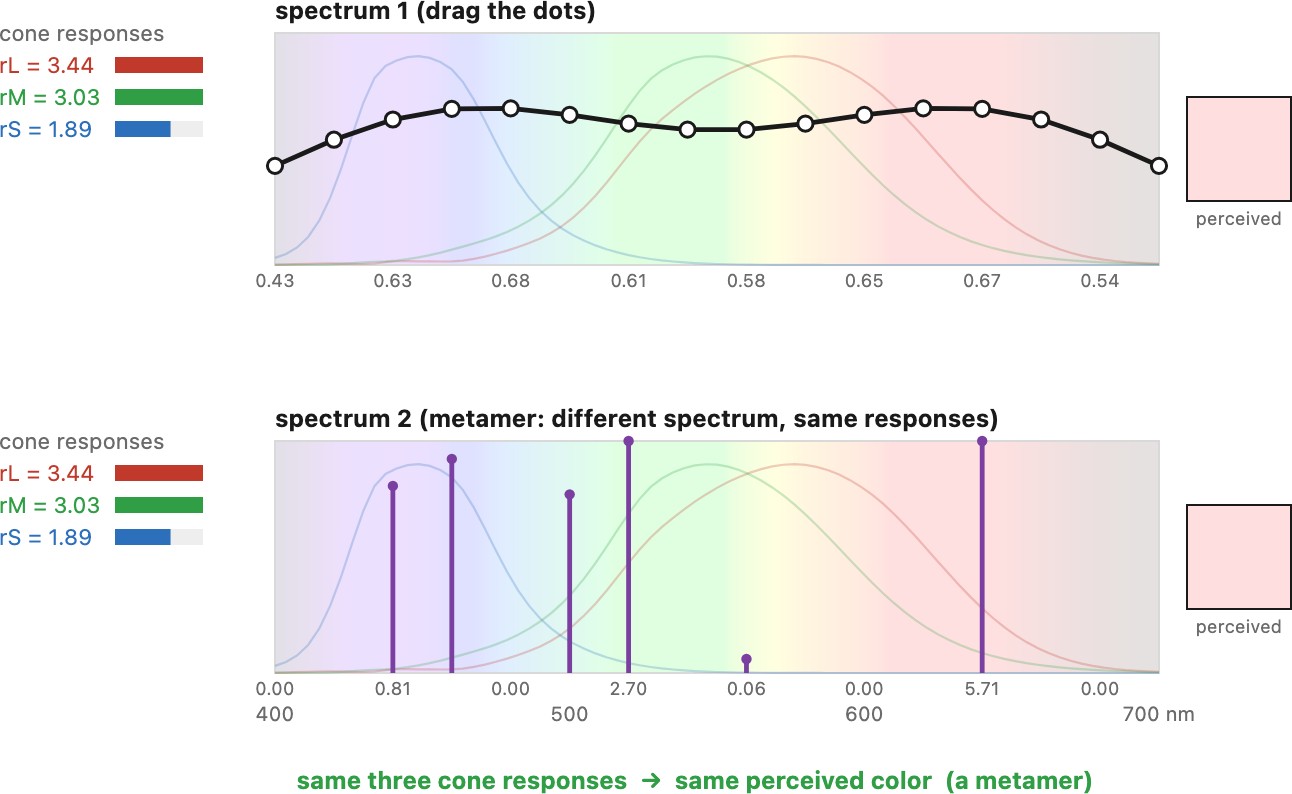
Because the map $\mathbf{E} \mapsto \mathbf{r}$ is a projection from infinite dimensions to three, it has an enormous null space: vastly many different spectra produce the same triple $(r_L, r_M, r_S)$ and therefore look exactly the same. Two such spectra are metamers. Metamerism is not a defect — it is what makes color reproduction possible at all. A screen with only three primaries can match (almost) any real spectrum, not by reproducing it, but by producing a metamer of it. The whole edifice of color technology in the next chapter rests on this.
Because color is only three dot products (L2.10), the map from spectrum to response has a huge null space: infinitely many different spectra collapse to the same triple and look identical — they are metamers. This is at once bad news — the eye throws away almost all spectral information, and two objects that match under one light need not be the same physically — and good news — you can reproduce any color with just three primaries, which is the only reason displays, prints, and the whole of color technology can exist.
Look again at the cone curves: the M ("green") sensitivity is entirely overlapped by L and S, so no real light can excite M without also exciting its neighbors. Every natural color is therefore a blend of cone activations, and the rainbow of nameable hues is exactly the set reachable by such blends. In 2025 a Berkeley team led by Ren Ng and Austin Roorda asked what lies outside that set. Their system, Oz (Fong, Doyle, Wang et al. 2025) uses adaptive optics to image the living retina and classify each cone by its type (L, M, or S — the technique of Roorda & Williams 1999), tracks the eye's constant fixational tremor at cellular scale, then delivers tiny laser microdoses to individually chosen cones — hundreds of thousands per second, each landing on the cone it was aimed at despite the moving eye. By addressing only the M cones at population scale it drives a pattern of cone activation that no spectrum of light could ever create, precisely because it sidesteps the cross-talk above. Observers report a color of impossible saturation, a blue-green they named "olo," so far outside the natural gamut that it can only be matched by conventional light after it is desaturated with added white. The lesson is sharp: the boundary of human color is not set by the eye's wiring but by the overlap of the cone curves — bypass the overlap by addressing cones individually, and you reach colors evolution never let us see. (It is the experimental mirror image of metamerism: metamerism is many spectra, one response; Oz is one response no spectrum can make.) Mechanically, Oz is the extreme of retinal projection — writing an image straight onto the photoreceptor mosaic — the frontier taken up in Retinal projection.

Most uses of metamerism save spectral information: three primaries stand in for a whole spectrum. Dolby 3D turns the idea sideways and uses it to separate two images that must look identical. A color 3D projector has to send a full-color picture to your left eye and a different full-color picture to your right eye off a single screen. Polarized systems separate the two channels by polarization; the old red/cyan anaglyph separates them by hue, at the cost of color. Dolby's system, the Infitec method of Jorke and Fritz (Jorke & Fritz 2003), separates them by wavelength. It splits each of red, green, and blue into two narrow bands — a "low" red near 629 nm for one eye and a "high" red near 615 nm for the other, and likewise for green and blue — and shows the left-eye image with one triplet of bands, the right-eye image with the other. The glasses carry matched interference (dichroic) filters: the left lens passes only the left triplet, the right lens only the right triplet, so each eye sees only its own image and the two stereo channels never cross. Metamerism is what makes the split invisible. The left eye's "red" (629 nm) and the right eye's "red" (615 nm) are physically different spectra, yet they are chosen and color-corrected so that both project to nearly the same three cone responses. They are engineered metamers. Both eyes therefore see the same red, the same full-color palette, even though the two eyes share no wavelength at all. This is the constructive side of metamerism (L2.11) pushed to its limit: not one spectrum standing in for another, but two deliberately non-overlapping spectra built to be color-identical, so one screen can feed two eyes at once. The price is in the glasses — precise multilayer dichroic coatings, far costlier than a sheet of polarizer or dyed plastic — plus a little light lost to the narrow bands, which is why you meet Dolby 3D in cinemas rather than living rooms.
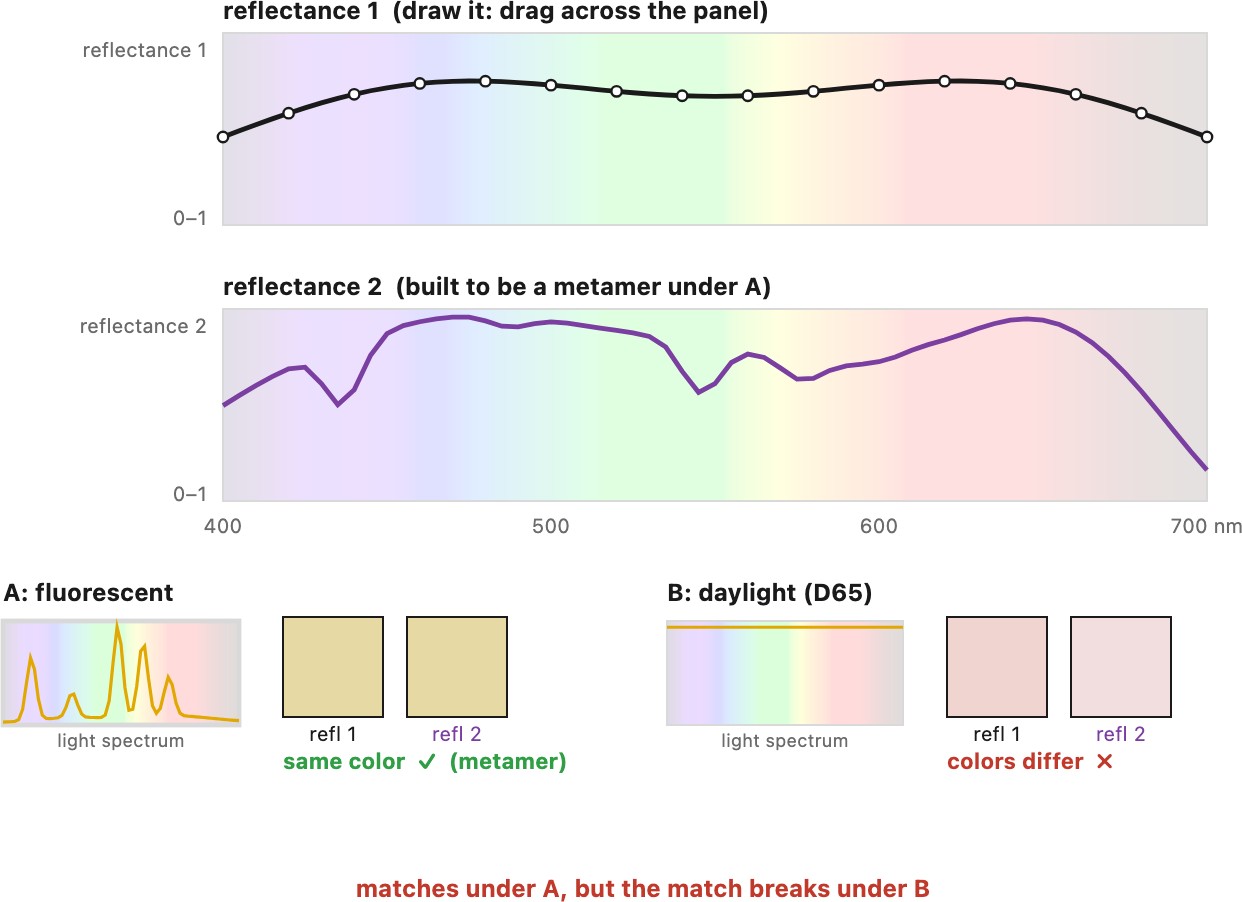
Metamerism also has an important limitation, and it follows from the very first lesson of this part. The light that reaches the eye is the product of a surface's reflectance and the illuminant (→ see Big lesson L2.6, illumination × reflectance), so a metameric match is really a match between two products. Change the illuminant — daylight for shop fluorescent, sunlight for tungsten — and you multiply both spectra by a different shape, and two reflectances that matched under one light can visibly diverge under another (Figure 2.2.11). The damage is worst under spiky illuminants (fluorescent tubes, white LEDs, sodium lamps), whose spectra are a few narrow peaks rather than a smooth curve. A spiky light samples the two reflectances only where its peaks fall, so small differences that a smooth daylight would average over get seized on and amplified, and the match falls apart. This is one of the situations where spiky illumination spectra are bad; we meet others under white balance and color rendering. The effect is easy to see for real (Figure 2.2.10): two gray patches printed as a metameric pair in Billmeyer & Saltzman's Principles of Color Technology look identical under daylight but split apart, one turning pink, under a warm halogen lamp. This illuminant metamerism is a real and expensive problem: a printer's inks are mixed to be a metamer of the target colors under the proofing light, then drift apart under the customer's living-room bulbs; and two pieces of an outfit, dyed to match in the shop, can clash in daylight. It is why color-critical work always specifies the viewing illuminant — "matches here" is never "matches everywhere."

A metameric match is a match between two products of reflectance and illuminant (L2.6). Swap the illuminant and you multiply by a different spectral shape, so two surfaces that looked identical under one light can diverge under another — illuminant metamerism. It is the headache behind prints that shift between the proofing booth and the living room, and fabrics that match in the shop but clash in daylight; color-critical work must always name the light a match is judged under.
In summary, going from physical color to the cone color is simple linear algebra. A linear projection or a big Nx3 matrix if you discretize wavelength. There is loss of information like with any projection, but the mathematical model is simple linear algebra. The part that makes the linear algebra messyare twofold. First, The three cone "axes" (the spectral responses) are not orthogonal — look again at Figure 2.2.4, where L and M overlap almost completely. Because $\mathbf c_L$ and $\mathbf c_M$ sit almost on top of each other across nearly the whole spectrum, their dot product $\mathbf c_L\!\cdot\mathbf c_M$ is large and positive: the two axes lean hard toward one another. In practice this means cross-talk — there is essentially no physical light that stimulates M without also stimulating L and S. Shine any single wavelength and at least two cone types respond together; the M signal can never be moved on its own — the only way to move it alone is to abandon light and stimulate the cones directly, cone by cone (the Oz sidebar above, and its extreme form in Retinal projection). Second, spectra, being physical power densities, cannot be negative, so the cone responses as well as any light we may use to reproduce color may not be negative. Both facts conspire against the comfortable situation in linear algebra where bases are orthonormal and each axis can be handled independently and where negative numbers are available to span the whole space.
The cone "axes" overlap heavily (L and M are nearly redundant) and light cannot go negative — and those two facts are what make color algebra genuinely awkward. With an orthonormal basis, finding a vector's coordinates is just projecting onto the axes; here it is not. Recovering the coordinates of a non-orthogonal basis requires a so-called dual (reciprocal) basis, and for a positive, overlapping basis the dual vectors point partly into the negative quadrants — the natural analysis directions have negative coordinates, which physical light can never supply. This is the deep reason there is no perfect set of primaries to reproduce all colors. (→ see Big lesson: Human color is non-orthogonal and non-negative.)

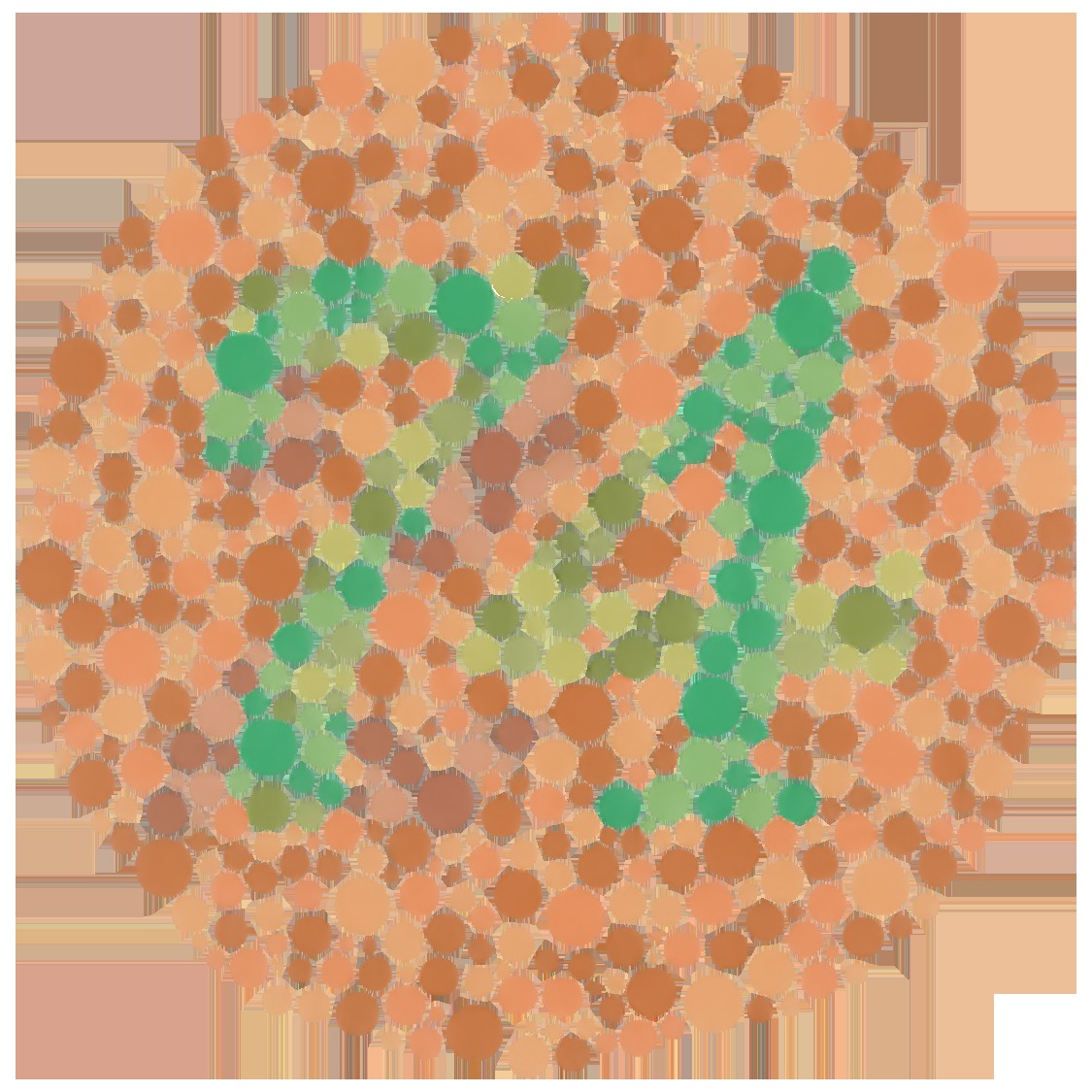
Color blindness is what happens when this projection loses even more information. A "dichromat" is missing one cone type (most commonly through the L/M genes), so the spectrum is projected onto two numbers instead of three; many spectra that look distinct to a trichromat now collapse into the same response — the classic red–green confusion. The standard Ishihara plates (Figure 2.2.13) exploit exactly this, hiding a figure that a trichromat sees and a dichromat cannot. The deeper point is one of perspective: a trichromat is also projecting away an infinity of spectral information. We are all color-blind relative to the full spectrum — and, as the next chapter (Animal eyes) shows, relative to other animals too.

Finally, the brain does not stop at three cone numbers. It re-codes color through a multistage model (simplified here; the full version is in Reinhard et al., Color Imaging):
- Stage 1 — cones. Three heavily overlapping L/M/S responses, the projection above.
- Stage 2 — opponent recoding, in the retina and the lateral geniculate nucleus. The L/M/S signals are re-mixed into three opponent channels: light–dark, red–green, and blue–yellow. This decorrelates the channels (since L and M are so redundant, their difference carries the color information efficiently) and matches the four "unique hues" we feel — the subject of the next section.
- Stage 3 — appearance. A higher-level organization closer to hue, saturation, and brightness (a hue-saturation-value-like, or HSV-like, arrangement), where context, adaptation, and memory shape the final percept — the domain of color-appearance models in the next chapter.
The recoding is not arbitrary. Color names cluster around shared focal colors across unrelated languages (Berlin & Kay's classic study, and the broader World Color Survey) — evidence that the structure is perceptual, built into the wiring rather than the culture. The recoding has support from two directions: psychophysics (Hurvich and Jameson's hue-cancellation experiments) and physiology (De Valois's recordings of opponent cells).
2.2.3 Opponent process and the multistage model⧉
The two-stage story is worth stating cleanly: color is trichromatic at the cones, then opponent in the retina and lateral geniculate nucleus. The three cone signals are recombined into three opponent channels — red–green (roughly $L - M$), blue–yellow (roughly $S - (L+M)$), and light–dark (roughly $L + M$, the luminance channel). In symbols this is just a fixed linear map,
with $\mathbf{M}$ a $3 \times 3$ matrix — the perceptual ancestor of the luma–chroma transforms (the $Y'C_bC_r$ of JPEG) we will use later (Figure 2.2.15).

Human color is not computed in one step. Stage 1 is trichromatic — three cone projections (L2.10). Stage 2 recombines them into opponent channels — roughly luminance ($L+M$), red–green ($L-M$), and blue–yellow ($S-(L+M)$) — that is, differences, not raw cone signals. This two-stage structure is why there is no "reddish-green," why staring at red leaves a green afterimage, why "primary color" is so confused (cones vs. opponent hues), and why luma/chroma encodings and chroma subsampling work — luminance carries the detail, the chroma differences are smooth.
This model earns its keep in two ways. It explains afterimages: stare at a saturated red patch until the red–green channel adapts, then look at white and the channel rebounds the other way, painting a green ghost (likewise blue→yellow). And it explains why luma/chroma encodings compress so well: the luminance channel ($Y'$, computed on gamma-encoded RGB — see Linear vs Gamma next chapter) carries most of the detail we can resolve, while the two chroma channels can be heavily subsampled with little visible loss — the foundation of JPEG's chroma subsampling, and a direct prediction of the opponent model that we will confirm under Spatial vision.
2.2.4 So what are the primary colors?⧉
Everyone has argued about this. Is it red-green-blue (RGB)? The grade-school red-yellow-blue (RYB)? Cyan-magenta-yellow (CMY)? The perennial muddle has two separate causes, and neither of them is a fact about light.
The first cause is the opponent recoding we just met. Because the visual system re-wires the three cone signals into red–green, blue–yellow, and light–dark, red, green, blue, and yellow all feel "primary" to us perceptually — there are four psychological unique hues, not three. This is why red-yellow-blue feels so natural to painters, and why the cones' actual L/M/S sensitivities match nobody's intuition of "primary" colors: our intuition is reporting the opponent stage, not the cones.
The second cause is additive versus subtractive synthesis, a matter of technology (taken up fully in the next chapter). RGB are the additive primaries — the colors of light you add together on a screen. CMY are the subtractive primaries — the inks or filters that remove light from white. The grade-school RYB is just a folk approximation of subtractive mixing.
So "primary color" silently conflates a perception fact with a technology choice — two different questions with two different answers, neither of which is intrinsic to the physics of light. As the lecture put it, the opponent rewiring "is one of the many causes of all the confusion about what's a primary color." (See the Glossary entry for primary.)
Recap: big lessons of this chapter
Each of the three cone types reports one number: the integral over wavelength of the incoming spectrum times that cone's spectral sensitivity — a projection (dot product) of the spectrum onto the cone curve, $r_k=\sum_i c_k(\lambda_i)\,E(\lambda_i)=\mathbf c_k\!\cdot\mathbf E$. Three cones → three dot products → the entire input to human color. Everything else about color — metamerism, "primary" colors, the color-matching matrices of the next chapter — follows from this one fact: color is three dot products with three fixed curves.
Because color is only three dot products (L2.10), the map from spectrum to response has a huge null space: infinitely many different spectra collapse to the same triple and look identical — they are metamers. This is at once bad news — the eye throws away almost all spectral information, and two objects that match under one light need not be the same physically — and good news — you can reproduce any color with just three primaries, which is the only reason displays, prints, and the whole of color technology can exist.
A metameric match is a match between two products of reflectance and illuminant (L2.6). Swap the illuminant and you multiply by a different spectral shape, so two surfaces that looked identical under one light can diverge under another — illuminant metamerism. It is the headache behind prints that shift between the proofing booth and the living room, and fabrics that match in the shop but clash in daylight; color-critical work must always name the light a match is judged under.
The cone "axes" overlap heavily (L and M are nearly redundant) and light cannot go negative — and those two facts are what make color algebra genuinely awkward. With an orthonormal basis, finding a vector's coordinates is just projecting onto the axes; here it is not. Recovering the coordinates of a non-orthogonal basis requires a so-called dual (reciprocal) basis, and for a positive, overlapping basis the dual vectors point partly into the negative quadrants — the natural analysis directions have negative coordinates, which physical light can never supply. This is the deep reason there is no perfect set of primaries to reproduce all colors. (→ see Big lesson: Human color is non-orthogonal and non-negative.)
Human color is not computed in one step. Stage 1 is trichromatic — three cone projections (L2.10). Stage 2 recombines them into opponent channels — roughly luminance ($L+M$), red–green ($L-M$), and blue–yellow ($S-(L+M)$) — that is, differences, not raw cone signals. This two-stage structure is why there is no "reddish-green," why staring at red leaves a green afterimage, why "primary color" is so confused (cones vs. opponent hues), and why luma/chroma encodings and chroma subsampling work — luminance carries the detail, the chroma differences are smooth.